Link to paper
The full paper is available here.
You can also find the paper on PapersWithCode here.
Abstract
- Neurons can emit action potentials in different patterns.
- Electrophysiologists labeled action potentials emitted at a high frequency as “bursts”.
- The burst coding hypothesis suggests that the neural code has three syllables: silences, spikes and bursts.
- Evidence is reviewed to support the ternary code in terms of mechanisms for burst generation, synaptic transmission and synaptic plasticity.
- Learning and attention theories are reviewed that benefit from the triad.
Paper Content
Introduction
- Problem of neural coding is to interpret neuronal signals
- Neurons represent input features with number of action potentials
- High impulse frequencies imply presence of feature
- Inputs are represented by computing averages
- Not all spikes are equal, code is ternary
- Evidence for burst coding hypothesis in 3 variations
- Alignment between generation and synaptic mechanisms
- Nervous system can process two streams of information simultaneously
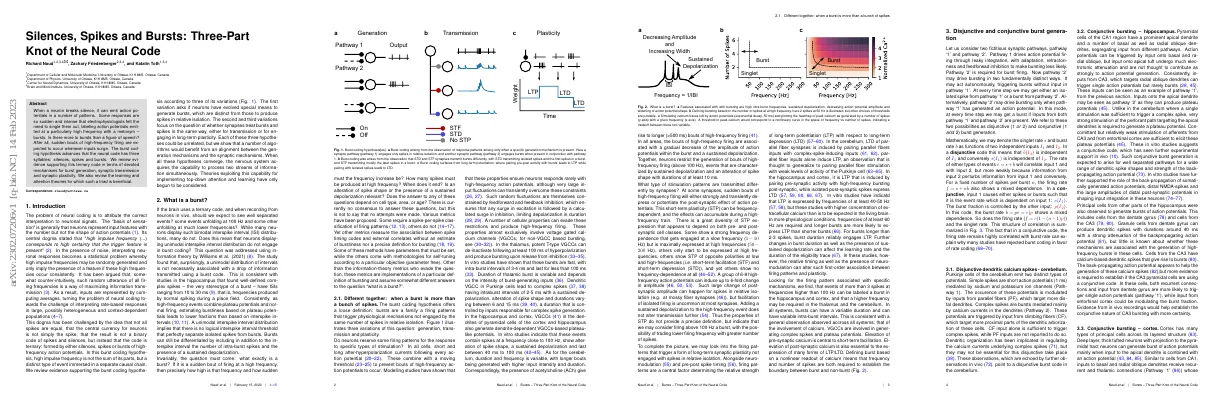
What is a burst?
- Brain may use a ternary code
- Neurons may display bimodal or unimodal interspike interval distributions
- A unimodal distribution of intervals is not necessarily associated with a drop of information transmitted using a burst code
- Burst coding hypothesis suggests that bursts are a family of firing patterns that trigger physiological mechanisms not engaged by the same number of spikes in relative isolation
- Features associated with bursting are high intra-burst frequencies, sustained depolarization, decreasing action potential amplitude and widening of action potential shape
- Defining bursting based on the number of spikes at a high frequency
- Synapses can show frequency-dependent short-term plasticity
- Firing patterns can trigger long-term synaptic plasticity
- Events of more than 3 spikes at frequencies higher than 100 Hz can be labeled a burst in the hippocampus and cortex
- A higher frequency may be required in the thalamus and the cerebellum
- Bursts can have a variable duration and can have variable intra-burst intervals
- Defining burst based on a nonlinear readout of calcium means that frequency and number of spikes are both required to establish the boundary between burst and non-burst
Disjunctive and conjunctive burst generation
- Pathway 1 and Pathway 2 are two fictitious synaptic pathways
- Pathway 1 drives action potential firing and Pathway 2 is required for burst firing
- Pathway 2 can act autonomously or drive bursting only when Pathway 1 has generated an action potential
- These two possibilities are referred to as disjunctive and conjunctive burst generation
- The rate of either types of events (spikes or bursts) is dependent on input 1 and input 2
- The firing rate is highly correlated with burst rate, which explains why many studies have rejected burst coding in favor of rate coding
Disjunctive dendritic calcium spikes -cerebellum.
- Purkinje cells emit two types of potentials: simple spikes and complex spikes
- Simple spikes are short action potentials mediated by sodium and potassium ion channels
- Complex spikes are bursts mediated mainly by calcium currents in the dendrites
- Complex spikes are triggered by input from climbing fibers, not parallel fibers
- Dendritic organization may regulate calcium currents underlying complex spikes
Conjunctive bursting -hippocampus.
- Pyramidal cells of the CA1 region have a prominent apical dendrite and a number of basal and radial oblique dendrites
- Input onto basal and radial oblique can trigger action potentials, but input onto apical tuft undergoes electrotonic attenuation
- Inputs from CA3 can trigger single action potentials but rarely bursts
- In vitro studies suggest a conjunctive code, which has seen further experimental support in vivo
- Backpropagating action potentials, distal NMDA-spikes and large amplitudes of distal post-synaptic potentials shape input integration in these neurons
- Granule cells from dentate gyrus and cells from CA3 can produce bursts of action potentials
Conjunctive bursting -cortex.
- Cortex has many types of principal cells
- Deep layer neurons can generate bursts of action potentials
- Inputs to basal and radial oblique dendrites receive recurrent and thalamic connections
- Bursting arises when one or both input pathways are active
- Correlation between singlet rate, burst rate, event rate, burst fraction, and firing rate with either inputs
- Cortex uses a conjunctive code where feedforward and feedback inputs are required to generate bursts
- Potent feedforward inhibition counteracts ability to generate action potentials
- T-type voltage-gated calcium channels control burstiness in thalamic relay neurons
- Controlling hyperpolarization controls burst fraction without affecting firing rate
Burst-dependent synaptic plasticity
- LTP is a cellular basis for learning and memory
- LTP requires NMDARs, elevated post-synaptic calcium and local glutamate release
- Calcium elevation at the synapse can come from NMDAR-mediated currents, local release from calcium stores or VGCC
- Post-synaptic bursting can control LTP expression
- Hebb’s theory needs to be revised when bursting is not triggered by the same pathway undergoing the potentiation
- In vitro recordings have confirmed a central role of post-synaptic bursting in the expression of LTP
- Plasticity in the cerebellum also shows burst dependence
- Burst-dependent learning rules in the electric sense of electric fish
Differential transmission of firing patterns
- Transmission must be frequency dependent to separate bursts from single spikes post-synaptically.
- Francis Crick hypothesized properties of synapses called von der Marlsburg synapses.
- STP is well established and can be triggered by burst-like events with a frequency closer to 100 Hz.
- STP can demix a mixture of spikes and bursts and can communicate independent information to different post-synaptic targets.
Cortex.
- Study surveyed over 20,000 synaptic pairs in animal slices and over 2500 in human tissue
- Most connections showed some level of STD over a wide range of stimulation frequencies
- Connections to and from PV-positive cells tend to show depression
- Some synapses show STD with a cutoff at 50 or 100 Hz
- Connections onto and from SST- and VIP-positive cells display STF
- Combining synaptic properties in cellular connectivity motifs can select bursts
- Polarization of local connections in terms of STD and STF is reflected in a polarization of impinging pathways
Hippocampus.
- CA1 pyramidal cells receive inputs from two main pathways: the perforant path and the schaffer collateral
- Perforant path inputs target mainly apical dendrites and NPY-positive cells
- Schaffer collateral inputs target mainly apical oblique dendrites, PV-, SST-as well as NPY-positive cells
- All inputs show short-term facilitation with frequency dependence
- CA1 pyramidal neurons project to multiple targets, with many connections going to the subiculum, which also display short-term facilitation
Cerebellum.
- Purkinje cells are the most numerous cells in the cerebellum.
- They receive inputs from granule cells and the inferior olive.
- PF inputs show STF, while CF inputs show an absence of short-term plasticity.
Thalamus.
- Sensory thalamus receives feedforward information from the senses.
- Thalamocortical afferents from sensory thalamus to sensory cortices show STD.
- Cortex sends feedback information to sensory thalamus with STF projections.
- Cortex sends feedforward information to higher-order thalamus with STD.
What logic.
- Inputs to principal cells are both STD and STF in the thalamus, more STD in the cortex and strictly STF in the hippocampus.
- Outputs from principal cells are strictly STD in the cerebellum and thalamus, but STF in the hippocampus.
- STD is used to communicate feedforward information, supported by the thalamus and cortex.
- STF inputs are used to control LTP in the cortex, hippocampus and cerebellum.
Algorithmic requirements for burst coding
- Representing information using two distinct types of activity can be beneficial for the nervous system
- Algorithms that require two streams of information can be implemented using a ternary code
- Supervised learning algorithms require two types of information
- Reinforcement learning algorithms benefit from burst coding
- Unsupervised learning algorithms require two types of information
- Attention signals take the form of gain modulation and plasticity gating, which can be facilitated by burst coding
Bursting in vivo
- Bursts occur spontaneously in awake and anesthetized animals
- Bursting is correlated with firing rate
- Different methods of measuring burst fractions lead to different results
- Anesthesia and sleep are associated with higher burstiness
- Relationship between arousal and bursting may extend to other areas
- Burst fraction in awake states is typically between 15-40%
- Variations in burstiness across areas indicate higher propensity in visual and prefrontal areas
Attention states.
- Visual attention enhances or suppresses saliency of locations and features in the visual field.
- Modulations of local field potential and noise correlations are observed in the visual cortex.
- Individual neurons show increases in firing rate and decreases in trial-to-trial variability.
- Direct measures of bursting modulation with attention have been observed in the visual cortex.
- When attention is directed onto a neuron’s receptive field, there is an increase in firing rate and a reduction in burstiness.
- In contrast, attention-dependent increases in bursting have been observed in anterior cingulate and prefrontal cortex.
- In the primary somatosensory cortex of rodents, changes in bursting correlate with a shift in perceptual performance.
Learning.
- Place fields are more likely to form in cells that produce bursts
- Correlation between spontaneous plateau potentials and place field formation
- Artificial induction of plateau potentials can give rise to a new place field
- Inputs from entorhinal cortex, CA3 and disinhibition from SST-positive are required for in vivo bursting
- EC bursting acts as an instructive signal
- High-frequency bursting of long duration occurs in cortex during associative learning tasks
- Blocking feedforward or feedback connections blocks learning
- Dendrite targeting inhibition also blocks learning
- Burstiness encodes errors with a delay of 1 second
- Silencing pyramidal cell activity during reward period alters learning of sensory association task
Discussion
- Brain uses a ternary code for representing, transmitting and processing information
- Presence of burst-generating calcium spikes depend on neuron size, cortical layer and species
- Neurons engage in bursting in vivo and have burst-dependent transmission and synaptic plasticity
- GABAergic interneurons of the cortex may show burst-dependent transmission
- Putative GABAergic cells modulate bursting in an attention task in vivo
Conclusion
- Ternary neural code is a way for cells to communicate both elevated membrane potential and elevated levels of intracellular calcium
- Synaptic plasticity is engaged by elevated calcium rather than elevated membrane potential
- Burst coding hypothesis suggests that synaptic pathways engage either spikes or bursts
- Burst coding also arises from the observation that STD and STF synapses transmit bursts differently
- Burst coding surfaces from long-term potentiation, pairing pre-post activity with bursts leads to LTP
- Features associated with bursting are high intra-burst frequencies, sustained depolarization, decreasing action potential amplitude and widening of action potential shape